
NOTE: A week or so before I posted this to the blog, John McPartland and Ernest Small’s new taxonomy for Cannabis was published without my having realized. It’s essential reading.
But there’s plenty here that’s still pertinent to understanding the ‘species controversy’, and getting why Cannabis comprises a single species, within which are two subspecies. There has never been any serious question whether there’s more than one species of Cannabis.
***
…sativa… indica… ruderalis… afghanica… americana… chinensis… gigantea…
Those are just a few of the names proposed for species—yes, species—of Cannabis.
But the prevailing consensus among botanists, universities, authoritative floras, botanical gardens, and national and international legislation is that Cannabis is a monospecific genus. In other words, most experts agree that the genus Cannabis is comprised of one highly varied species, C. sativa L.
Proponents of multispecies classifications fight on—most popularly, Mark Merlin and Robert Clarke, who have proposed a classification based on just four studies by Karl Hillig. No central authority or academy presides over the discipline of taxonomy. The discipline itself is both a theoretical science and a practical art. But it’s the “lumpers” who continue to prevail over the “splitters” in the Cannabis species debate.
As Ettiene DeMeijer puts it: “A monospecific concept… has implicitly been adopted in virtually all nontaxonomic publications on Cannabis… The current pattern of Cannabis diversity is primarily due to intentional actions of humans and reflects a long, intense, and divergent process of domestication which has blurred any natural evolutionary pattern of diversity. It is even questionable if truly wild Cannabis still exists.”
Questionable is an understatement. No truly wild populations of Cannabis have ever been identified, and it’s unlikely any ever will be.
“No persuasive evidence has been documented that there are truly wild populations of C. sativa (pristine genetically, never having been altered by human selection, and having natural distributions)”, writes Ernest Small, adding that “wild-growing plants of Cannabis sativa, insofar as has been determined, are either escapes from domesticated forms or the results of thousands of years of widespread genetic exchange with domesticated plants, making it virtually impossible to determine if unaltered primeval or ancestral populations still exist. Moreover, because the species has been spread and modified by humans for millennia, there does not seem to be a reliable means of accurately determining its original geographical range [though see McPartland] or even whether a plant collected in nature represents a primeval wild type or has been influenced by domestication.”
In other words, Cannabis is a cultigen, in the sense of a crop for which there is no known wild ancestor. There are hundreds of other examples, including the avocado, aubergine, onion, and peanut. In its broadest sense, the term cultigen refers to a plant that’s resulted from artificial selection. Fundamentally, Cannabis is a creation of humankind—a domesticate.

Ernest Small in his field of Cannabis, grown on behalf of the Canadian government at the Central Experimental Farm in Ottawa in 1971.
The best taxonomy for Cannabis proposed to date, namely Small & Cronquist, recognizes this central characteristic. In Cannabis, the essential pattern of diversity is the creation of humans selecting for desired traits. The main divergence is between domesticates with high levels of stem fibre and domesticates with THC-rich inflorescences. So, we have a single species, C. sativa L., comprised of two subspecies, subsp. sativa (“hemp”) and subsp. indica (“marijuana”), which have diverged due to artificial selection.
Because hemp and marijuana are creatures of human creation to classify them as two distinct species (as in Clarke & Merlin or Schultes) is to commit an error according to taxonomic theory – specifically with respect to how formal classification applies to domesticates. As Small writes, “the recognition of more than one species of Cannabis is typical of the overclassification of domesticated crops.” This is a mistake often made by botanists who aren’t trained taxonomists.
For practical purposes, in Small & Cronquist, subsp. indica and sativa are divided on the basis of 0.3% THC, a utilitarian criterion which most countries have adopted for legislative purposes—though Small himself has said that this number needs to be reviewed. Again, taxonomy is both a theoretical science and a practical art.
The Small & Cronquist system is supported by genetic research:
Mandolino et al. (2002) quantified DNA polymorphisms in ten drug- and fiber-type varieties and identified more variability between individuals within a varietal than between varietals—data that confirmed “the existence of a single, widely shared gene pool.” Gilmore et al. (2007) analyzed a globe-spanning collection and found a low rate of sequence variation, which is consistent with a single species. McPartland and Guy (2018) performed a DNA barcode analysis of pairs of plant species, subspecies, and varieties. Subsp. indica and sativa diverged with a mean barcode gap of 0.41%. This was very close to the mean of 0.43% for other plants at the rank of subspecies or variety such as small-leaved and large-leaved tea (Camellia sinensis var. sinensis and C. sinensis var. assamica.)
All of this points to Cannabis being a single species with two subspecies.

Classification of Cannabis sativa L. by Small & Cronquist (1976), illustrating conceptual bases of delimitation. Source: Small (2015)
This brings us to the next rank down in the Small & Cronquist taxonomy. Here, subsp. indica and sativa are each split into two subgroups, namely their respective cultivated (“farmed”) and uncultivated (“wild” or weedy) varieties. For Cannabis aficionados, it’s important to understand that there are two senses of the term variety: the looser more colloquial sense often applied by gardeners versus the strict botanical term employed by taxonomists. Horticultural and agricultural varietals or varieties are named using English “fancy names”, which are put in quotation marks—for example, ‘Cherry Parfait’ or ‘Falstaff’. Notional equivalents in Cannabis would be ‘Northern Lights’ or ‘Green Crack’. But when wielded formally in taxonomy, the word variety refers strictly to the rank immediately below subspecies and involves Latinized nomenclature. This results in a straightforward four-part system.
- C. sativa subsp. indica var. indica – e.g. Asian drug-type landraces from regions such as Thailand and Afghanistan, hybrids such as Skunk and Blueberry, and registered hybrid cultivars, whether they’re types known colloquially as Sativas or Indicas.
- C. sativa subsp. indica var. kafiristanica – e.g. wild-type populations from regions such as the Himalaya and northern India.
- C. sativa subsp. sativa var. sativa – e.g. Chinese hemp landraces such as Liuan Huoma and European cultivars such as Finola.
- C. sativa subsp. sativa var. spontanea – e.g. wild-type populations from regions such as eastern Europe and eastern North America.
Achenes (“seeds”) are the main means used to differentiate domesticated from wild-type varieties of Cannabis, primarily based on retention or shattering. Wild-type inflorescences readily drop their seeds, such that at the end of summer few if any seeds will remain on a plant. Cultivated types retain their seeds within the inflorescence. In addition, wild-type seeds are small and typically mottled with elongated bases and an abcission zone that facilitates shedding, whereas landraces and modern cultivars have markedly larger seeds, relatively minimal mottling if any, no extended base, and a minimal abcission zone. Respectively, these are called the wild and domesticated seed syndromes. They’re divergent traits that result from disruptive selection for existence under cultivation or in the wild.
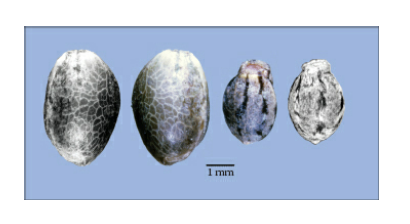
Achenes (“seeds”) of C. sativa (areas of attachment to the plant are uppermost). Left side shows two achenes of a domesticated plant, right side shows two achenes of a ruderal plant. The domesticated fruits are larger, lack a camouflagic persistent covering layer derived from the perianth, and lack an elongated attachment base that facilitates disarticulation in the wild form.
Small, Ernest. Cannabis: A Complete Guide.
But leaves matter here too. Cannabis has compound leaves. Like other crops with compound leaves, such as the carrot or alfalfa, cultivated Cannabis has large leaves, whereas its wild-type populations have small leaves. Large leaves here means those with a large surface area, whether caused by leaflets that are long or broad or both. These characteristics are adaptive. Large leaves with wide leaflets photosynthesize more efficiently. By contrast, small leaves and narrow leaflets have survival value outside of cultivation. They’re hardy traits. Evidence suggests, for example, that narrow leaflet width can hinder grazing by insects.

‘Deep Chunk’, an heirloom Afghan inbred from a ’70s landrace accession by Tom Hill
The significance of large or small leaves relates to whether we can or should formally classify so-called “Indicas” and “Sativas”. Formal classification involves using Latinized names. Close to all formal biological classification is applied to wild (i.e., undomesticated) organisms. But Indicas and Sativas are cultigens in the narrow sense of groups that are distinguishable by attributes that have been differentially selected by humans. Hillig and Clarke refer to them as “Broad Leaf Drug” (BLD) and “Narrow Leaf Drug” (NLD) biotypes (the accurate term here would be leaflet, not leaf). Small uses “Cannabis Group Narcotic, THC/CBD Balanced” and “Cannabis Group Narcotic, THC Predominant”. Leaving aside whether these categories have value, there’s nothing wrong per se with labelling these two types of “marijuana” cultigen using informal (i.e., non-Latinized) nomenclature. The problem arises when attempts are made to recognize them formally as taxonomic groups (i.e., “taxa” such as species, subspecies, varieties). The trouble with the two taxa Cannabis indica subsp. afghanica and Cannabis indica subsp. indica proposed by Clarke and Merlin is they they ignore traditions and theories of how variants of domesticated creatures such as Cannabis should be classified.

")
What Clarke and Merlin do with subsp. indica and afghanica is further split Schultes’ (Schultes et al., 1974) splitter concept of Cannabis indica.
Until 1971, Schultes himself was an eloquent advocate for lumper orthodoxy. As he wrote in 1970: “Botanists now generally agree that Cannabis is a monotypic genus, a genus with one species, C. sativa, that there cannot be recognized any true botanical varieties within this species, and that this one species has diversified into a great number of ecotypes and cultivated races. Modern taxonomists, thus, are in agreement with Linnaeus’ treatment of the genus.” But following a 1971 trip to Kandahar instigated by an enterprising lawyer for the Brotherhood of Eternal Love, Schultes had a sudden change of heart. He proposed his three-species C. ruderalis, C. sativa, and C. indica system, in which C. indica is comprised of all Afghan, Indian, Middle Eastern and other “intoxicating” types. By the late ’80s, it appears he may have come round to Anderson’s concepts of reserving C. indica exclusively for dwarf Afghan types and making C. sativa an incongruous combination of fibre hemp and narrow-leafleted marijuana cultigens (Anderson, 1980; Schultes and Hoffmann, 1987). Out of this chaos was born the “vernacular taxonomy” of Indica–Sativa–Ruderalis. Afghanica has since been added to the confusion.

Anderson’s three-species taxonomy, foreruner of the vernacular taxonomy of Indica, Sativa, and Ruderalis
Clarke and Merlin rationalize their proposed subsp. indica versus afghanica split based on their interpretation of studies by Nikolai Vavilov and Karl Hillig. But typological thinking appears to have led to their misreading Vavilov.
[NOTE: the tangled issues addressed in the following paragraphs appear to have been resolved by McPartland and Small’s 2020 study, ‘A classification of endangered high-THC cannabis (Cannabis sativa subsp. indica) domesticates and their wild relatives‘, which establishes an apparent naturally differentiated pattern of variation within wild-growing populations of subs. indica, namely between its Central and South Asian centres.]
In 1924, Vavilov undertook an extensive expedition across Eurasia that included Afghanistan. His interpretations of what he observed are often protean and contradictory. But the picture he had arrived at by 1929 is simple. He perceived two main Eurasian populations, northern (C. sativa) and southern (C. indica). The Kunar Valley and the mountains of the Northwest Frontier of British India formed a dividing line between them. Indian cultigens (Cannabis indica Lam.) were derived from their own independent centre of origin—the wild populations of Kunar Valley and the mountains to its south and east in what is now Pakistan. To the north of Kunar, Vavilov observed hashish production, most of it in regions of northern Afghanistan such as Balkh, Badakshan, and Herat. He characterized these hashish landraces as Turkestani, like those of Bukhara and Samarkand in Central Asia proper. Their leaves were large, with long leaflets, for which he provides length measurements. But their leaflets appear to have been narrow, or relatively so, because in his comparitive tables the only landraces Vavilov notes as having definitively broad leaflets were those he observed in China. [Edit: my inference was mostly incorrect, see new McPartland & Small herbarium study] In his 1920s treatments, he classified both Chinese hemp and these Turkestani hashish plants as C. sativa L. In a later treatment (published 1949–51) he appears to have moved Turkestani hashish landraces to C. indica. Regardless, all of this begs the question of where in Vavilov’s work to find the dwarf broad-leafleted cultigens we associate with the Hindu Kush—whether we call them Indicas or Afghanicas.
Clarke and Merlin believe they have found them in Vavilov’s account of Kunar. The relevant section from Cannabis: Evolution and Ethnobotany is worth quoting at length:
“Vavilov and Bukinich (1929) first reported the occurrence of the Afghan C. indica broad-leaf drug (BLD) biotype recognized by Hillig (2005a). […] Although Vavilov never reported observing NLD varieties in South Asia, he included two BLD varieties from Afghanistan and Pakistan as subspecies of C. indica. Because the Afghan varieties differed greatly in gross phenotype and biochemistry from familiar European NLH varieties of C. sativa but were native to the Indian subcontinent, Vavilov assigned them to the second established taxa, C. indica, rather than to C. sativa. Vavilov did not mention if the Afghan varieties were drug or fiber types, although based on later observations from Afghanistan (Schultes et al. 1974) and neighboring regions, we suggest that they were used almost exclusively for hashish production. Although Vavilov included BLD varieties from Afghanistan within C. indica, he assigned them to a subspecies rank of afghanica (Vavilov and Bukinich 1929) since they were from Afghanistan, not India, and were dissimilar to Lamarck’s NLD-type specimen in gross phenotype, leaf shape, and inflorescence morphology. Vavilov and Bukinich’s nomenclature is also supported by Hillig (2004, 2005a/b) and Hillig and Mahlberg (2004). In 1972, Richard Evans Schultes of Harvard University traveled to Afghanistan specifically to obtain specimens of then little-known Afghan hashish cultivars. Following Vavilov, Schultes accepted a broad interpretation of C. indica to include Afghan varieties, as well as Indian drug types and their new world descendants, much as we presently accept. Therefore, we now include the short, broad-leaved, acrid smelling Afghan Cannabis in the BLD biotype (C. indica ssp. afghanica), as described by Vavilov and Bukinich (1929) and Schultes et al. (1974) and presently referred to as the “indica” or “hashish plant” by domestic marijuana growers.”
This significantly misrepresents Vavilov’s accounts. He does not describe the populations of Kunar as broad-leafleted. In fact, he states in the body of the text and in tables that their leaflets have a “narrow, obovate shape”. He added that this trait “was not observed by us among the European, Siberian and Turkestani forms.” And while it is true that “Vavilov did not mention if the Afghan varieties were drug or fiber types”, this phrase obscures that he explicitly stated he believed Kunar Valley to be a region where Cannabis “is definitely not cultivated”, noting that “Wild [Cannabis] has not been encountered in any other areas of Afghanistan.”

Vavilov’s type specimen for his proposed taxon Cannabis indica var. kafiristanica. In all respects except its seeds (white, with domestication syndrome), f. afghanica was identical. For the true signficance of this specimen, see the crucial new study by McPartland and Small (April 3rd 2020).
Vavilov’s concept of the afghanica taxon was markedly different from that proposed by Clarke and Hillig. Initially, in 1924, he appears to have thought of the Kunar population as comprising wild-type relatives of Turkestani landraces, and assigned it to C. sativa var. afghanica, akin to his C. sativa var. spontanea taxon for ruderal hemp, though he published no type specimen. In 1927, after cultivating seeds from a Kunar accession, he revised his concept. Now, he moved the population to C. indica Lam. In 1929, he published a photograph of a specimen of C. indica var. kafiristanica. This was a very young plant cut from a crop grown in St. Petersburg (60° N) in Russia, which due to age and phenotypic plasticity makes placing weight on morphological characteristics such as its branching and leaflet shape problematic. In 1929, he made further revisions, splitting the Kunar population into C. indica var. kafiristanica and C. indica f. afghanica. Form (f.) is a rank below variety; for example, white-flowered plants of a species that typically has coloured flowers may be classified as f. alba. The only feature of f. afghanica that distinguished it from var. kafiristanica was its seeds, which were light coloured with domestication traits. “In all its other characters, it resembles the typical wild eastern Afghan [Cannabis].” For Vavilov, f. afghanica was an intermediate form connecting the wild populations of the eastern Hindu Kush and northern Pakistan to the Lamarckian ganja cultigens of India. (“In the characters of its fruit, it seems to be a link between the typical wild forms and the cultivated forms.”) The mountain frontier of what was then British India was for Vavilov a dividing line beyond which Cannabis “is represented by an independent focus of form-derivation, apparently being a special Linnean species.” Of the typical Afghan hashish landrace he had observed, he wrote, “In all its features it is directly related to the Central Asian cultured [Cannabis] and not to Cannabis indica Lam., which is distinguished by small leaves, small fruit, and low growth (not more than 1 metre).” He added, “The wild East-Afghan [Cannabis] (var. kafiristanica Vav.) is conversely, related to the cultured C. indica Lam. cultivated in India.”

‘Gathering the Ganja Crop’ in the Ganja Mahal, Naogaon, East Bengal, now Bangladesh. A photo from the Indian Hemp Drugs Commission (1894).
We now know that Vavilov had mistaken notions about Indian landraces, such as that they’re necessarily short. They aren’t, and in so far as they appear to be, this is with few if any exceptions related to phenotypic plasticity and cultivation techniques. Vavilov’s misconceptions informed Schultes’ 1974 splitter taxonomy. Now, it may be that Clarke and Hillig are onto something, and there were cultivated broad-leafleted populations in or around Kunar. I discuss that likelihood here on the Curious About Cannabis podcast. But the morphology of a very young ruderal plant grown out in St. Petersburg (at a similar latitude to Alaska) is at best a tenuous basis for claiming that Vavilov himself observed BLD Afghans. Many narrow-leafleted cultigens, such as those from the Himalaya or India, can exhibit markedly broad leaflets during early vegetative growth. Moreover, the most accurate description of Vavilov’s type specimen’s leaflets is oblanceolate (i.e., “obovate, but narrow”). Similar problems of lack of adequate accessions undermine Hillig’s four studies. Of 135 accessions, only 11 originate from in or around the Hindu Kush. Of these, only three or four are direct from Afghanistan or Pakistan. The others are from Amsterdam and appear to be inbred cultivars rather than actual landraces. In addition, there is one ruderal accession from Uzbekistan.
Where Hillig’s studies have value is in identifying genetic relationships. But it’s essential to grasp that with a creature of domestication such as Cannabis, conclusions concerning heredity are not straightforwardly transferable to taxonomy. Hillig (2005a,b) has applied multivariate statistical similarities of allozyme frequency to differentiate European fibre plants from the three eastern domesticated groups, namely the two marijuana cultigens (Indicas and Sativas) and Chinese fibre plants. Weaker support for this divergence was provided by terpene chemistry (Hillig 2004a) and cannabinoid chemistry (Hillig and Mahlberg 2004), and the evidence was clearer for cultivated accessions than for ruderals. Hillig assigned European fibre plants to C. sativa and the three eastern groups to C. indica. This combined within C. indica the two marijuana groups and Chinese hemp. Hillig’s data indicate an ancient trend of genetic differentiation between the plants of western Eurasia (and consequently Europe) and those of eastern Eurasia. European hemp appears to have undergone a genetic bottleneck when being selected from eastern populations. McPartland examined pollen data which could support this, suggesting widespread Cannabis cultivation was introduced to eastern Europe by the Scythians in the late Bronze and early Iron Ages.
But Ernest Small comments as follows:
“…by evolutionary standards, this trend seems very minor, since not a single reliable character has been found to distinguish the western (European) and eastern kinds collectively, nor has a combination of morphological characters been suggested that could serve to separate them reliably, as is necessary in conventional plant taxonomic identification keys. Recent DNA evidence does indicate that at the molecular level, combined genetic loci may be usable to discriminate European hemp strains, Indica type plants, and Sativa type plants (Lynch et al. 2015; Sawler et al. 2015). The situation is perhaps analogous to human blood group geography, thought to have resulted from a combination of random drift and selection for disease resistance (Anstee 2010), and certainly not warranting formal taxonomic recognition. The information is, however, useful for tracing genetic relationships and identifying strains and cultivars.”
Genomics is generating great excitement in the Cannabis field for good reason. But caution is advised before applying genetic data to taxonomy. There continues to be a paucity of accessions, particularly of subsp. indica landraces and wild-type populations. And equally, those of us who are not well-versed in the traditions and theory of taxonomy—which is most of us, and I include myself in that—are poorly placed to interpret results. Fundamentally, this relates to the problem of how a domesticated creature such as Cannabis can or should be categorized and how to understand proposed classifications and labels for its variants. The use of genealogical (genetic) relationships for classification of true wild groups is firmly established. But crucial theoretical concerns persist regarding how to apply them to domesticated groups.

Pingback: McPartland & Small Publish New Cannabis Taxonomy | The Real Seed Company Blog·
Pingback: Landrace Interview with Angus on ‘Curious About Cannabis’ | The Real Seed Company Blog·
Pingback: ‘Curious About Cannabis’ Landrace Interview on Youtube | The Real Seed Company Blog·
Pingback: Endangered Varieties of subsp. indica: A Few Thoughts | The Real Seed Company: The Honest Online Source for Cannabis Landraces Since 2007·
These articles are so educational and interesting. Thank you for making this information available.
Peace farmerlion
MedDakotabis
thanks, glad to hear it!
Great job on compiling the information and overview. It’s a very interesting and debated subject.
This article saves a lot of motion and effort from the reader by having it all summed up and digested in one place. You’re a gem Angus.
Much respect and thank you for your contributions.
Hope to see you on ICMAG, I do miss your posts.
Thanks, much appreciated. It is indeed an interesting subject. There’s not much left to debate, however. Nor was there ever much, to be fair. On the one side we have the taxonomists and evidence, on the other, a handful of folks either acting in bad faith for good reasons (Schultes) or without a solid grasp of taxonomy or the field (Clarke etc).
Appreciate your remarks about icmag etc but not a chance!
Pingback: What’s the Real Meaning of ‘Landrace’? | The Real Seed Company: The Honest Online Source for Cannabis Landraces Est. 2007·